
谈糖不色变
岩藻糖与麦角硫因
在细胞稳态与抗衰老中的最新研究
2025年09月25日 16:59

衰老被认为是由多因素驱动的复杂过程,具体可参考《Cell》2025年提出的衰老的"十四大标志" (Kroemer & Maier, 2025)。与衰老相对应的,《Cell》也在2023年提出了健康的“三大标志”,其中包括了稳态维持以及对应激的适当应答。衰老的诸多标志也确实表现为细胞稳态的逐渐丧失以及对应激的应答能力下降(Lopez-Otin & Blasco, 2023)。因此,从细胞稳态和对应激的应答能力入手,开发具有干预潜力的活性物质成为近年来抗衰研究的重要方向。
岩藻糖和麦角硫因作为天然来源的小分子,分别作用于蛋白折叠和线粒体,在维持细胞稳态方面显示出独特优势。本文综述两者的作用机制,并探讨其联合应用在护肤抗衰领域的潜在价值。


线粒体是细胞能量代谢的中心,其功能衰退被视为衰老的"十四大标志"之一 (Kroemer & Maier, 2025)。线粒体DNA突变、蛋白稳态缺陷导致呼吸链复合物不稳定、线粒体呼吸传递链(ETC)效率下降、线粒体数量的减少以及线粒体膜电位的异常等,都会导致线粒体的呼吸效率下降,具体表现为耗氧量减少以及耗氧转化为ATP效率下降,加速细胞老化(Lopez-Otin & Blasco, 2023)。如果对线粒体的异常不进行干预,细胞会陷入能量合成不足与 ROS(reactive oxygen species)过度产生形成恶性循环,导致线粒体的降解和破坏,开启细胞的程序性死亡 。

蛋白质承担着结构支撑、信号转导、酶促反应和代谢调控等几乎所有细胞生物学功能,这些功能依赖于蛋白质的正确折叠和动态更新。因此,蛋白质稳态的丧失同样被视为衰老的"十四大标志"之一 (Kroemer & Maier, 2025)。衰老及多种与年龄相关的疾病,如肌萎缩侧索硬化症(ALS)、阿尔茨海默病、帕金森病和白内障,均与蛋白稳态受损相关(Lopez-Otin & Blasco, 2023)。蛋白稳态失衡会导致蛋白聚集和内质网应激损伤,从而加速衰老和疾病。而增强蛋白稳态可以提高蛋白折叠/降解能力,延缓衰老并改善健康寿命。未折叠蛋白反应(UPR)是蛋白稳态与延缓衰老的重要联系节点。而分子伴侣蛋白(ER chaperone)是UPR通路的主要效应因子之一。
• 蛋白稳态和内质网
内质网(ER)不仅是脂质与蛋白质合成的关键场所,也是细胞信号网络的重要枢纽(Walter & Ron, 2011),与线粒体等细胞器在物理和功能上形成动态网络,以协调蛋白质折叠、脂质代谢、钙稳态及细胞应激反应等核心过程。蛋白错误折叠会激活内质网的应激反应,从而引发未折叠蛋白反应(UPR)。UPR可以恢复蛋白稳态,清除错误折叠蛋白,但在应激无法缓解的情形下,也可能诱导细胞凋亡(图1)。
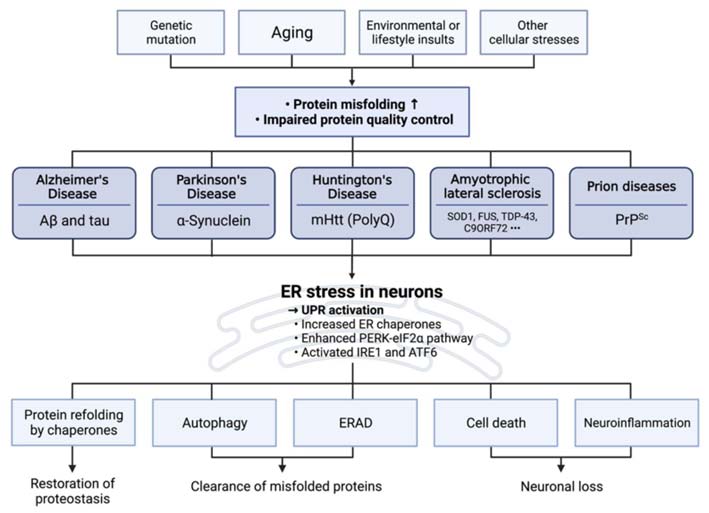
图 1 内质网应激及其关联系统与衰老的关系
• 未折叠蛋白反应(UPR)和蛋白稳态
当未折叠或错误折叠的蛋白在内质网腔内积累时,细胞会激活UPR以恢复蛋白质稳态(Ron & Walter, 2007)。UPR主要通过三方面发挥作用:(1)下调蛋白合成并同时增强折叠能力,通过减少胞内的未折叠蛋白数量来减轻内质网应激;(2)通过内质网相关降解(ERAD)与自噬途径,清除错误折叠蛋白;(3)调控细胞命运,当应激不可缓解时,UPR可诱导细胞凋亡(Hetz, 2012)。
• 分子伴侣蛋白和蛋白稳态
分子伴侣蛋白(ER chaperone)是UPR通路的主要效应因子之一。内质网分子伴侣蛋白在保证内质网内蛋白质折叠质量和内质网稳态上,发挥了重要作用。分子伴侣可以与新合成蛋白的疏水区或未折叠区结合,防止它们错误折叠或聚集。它还可以识别未折叠或错误折叠的蛋白,将无法折叠的蛋白送往内质网相关降解(ERAD)通路降解,避免毒性聚集。当细胞内未折叠蛋白积累时,分子伴侣基因表达上调,从而增加折叠能力、降低错误折叠蛋白的积累。

内质网和线粒体并非孤立工作,它们通过高度动态的物理和功能连接——线粒体相关内质网膜(MAM) 紧密协作,协调Ca²⁺信号、能量代谢与应激应答,共同调控细胞稳态。由于内质网与线粒体等细胞器在物理和功能上动态网络的存在,蛋白质错误折叠及其聚集不仅直接损害内质网功能,也会通过破坏线粒体稳态和诱导氧化应激,进一步加剧细胞衰老(Lopez-Otin & Blasco, 2023)。图2展示了从蛋白折叠错误最终引发线粒体功能异常和细胞凋亡的路径。内质网和线粒体之间的信号传递包括了Ca²⁺和ROS等。
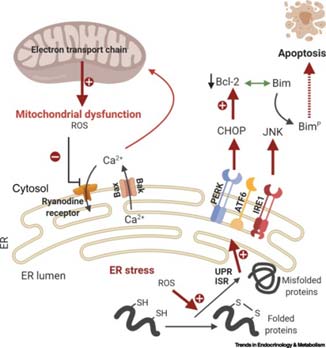
图 2 从蛋白折叠错误到线粒体功能异常

糖基化(包括岩藻糖基化)是蛋白最常见的翻译后修饰之一,指的是将各种糖基共价连接到多肽链上。蛋白上的糖基在糖蛋白折叠及糖基介导的分子相互作用中具有关键作用,其功能取决于糖基的具体结构。
研究发现(Wang & Tan, 2023),FUT2(其表达会被岩藻糖上调)介导的岩藻糖基化通过修饰并稳定内质网分子伴侣蛋白HYOU1,维持蛋白质折叠质量控制和UPR平衡,还可以抑制 UPR 的促凋亡分支(IRE1/TRAF2/ASK1/JNK 信号)(图4),减少氧化应激和线粒体相关的凋亡。当岩藻糖基化不足时,HYOU1稳定性下降,内质网应激增强,UPR反应失衡并诱导细胞凋亡。这揭示了岩藻糖和岩藻糖基化在”内质网稳态—UPR—蛋白折叠“路径中的关键作用,也为岩藻糖在细胞保护和抗衰老中的应用提供了分子机制依据。从图3可以看到,岩藻糖通过”内质网稳态—UPR—蛋白折叠“路径,显著提升了因LPS诱导丧失的线粒体膜电位(MMP),改善了受损的线粒体功能,抑制了与线粒体途径密切相关的凋亡因子BAX、CYTC和活化的Caspase-3的表达,提升了抗凋亡因子BCL2的表达。改变了细胞凋亡的命运。
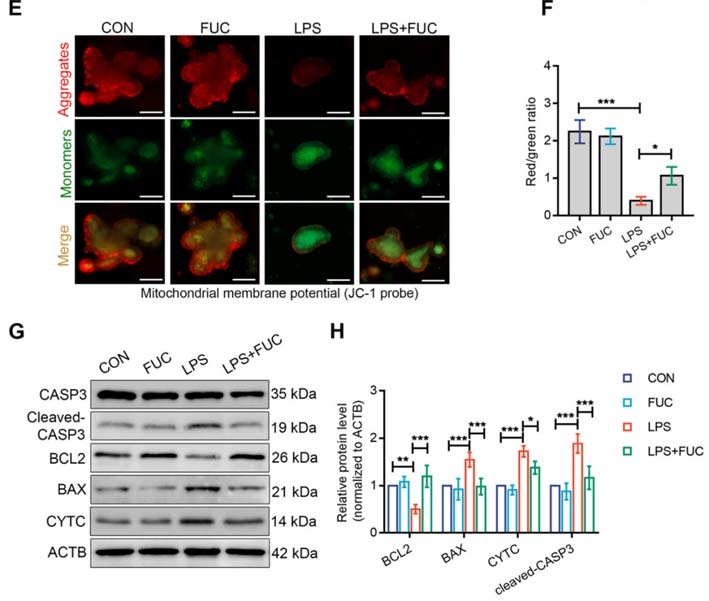
图3 岩藻糖通过内质网稳态—UPR—蛋白折叠轴影响线粒体功能和细胞命运
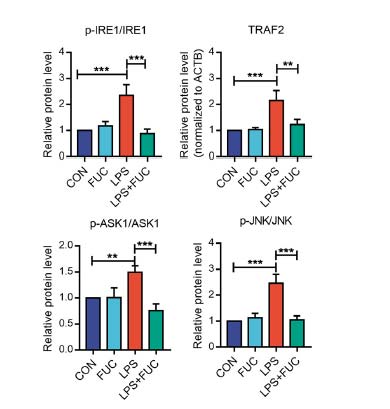
图4 岩藻糖抑制 UPR 的促凋亡分支因子表达(IRE1/TRAF2/ASK1/JNK)
华熙生物在皮肤相关细胞上做的结果也发现,岩藻糖可能通过这条”内质网稳态—UPR—蛋白折叠“路径,既大幅降低了 LPS 诱导的成纤维细胞 ROS,极大缓解了细胞内的氧化应激(图5),又恢复了 UV 损伤下角质形成细胞的屏障蛋白表达(图7),并增强其迁移修复能力(图6),同时在免疫细胞中减弱了 LPS 驱动的促炎因子 IL-6 表达(图8)。这些结果共同揭示了岩藻糖在细胞保护、抗炎和组织修复中的关键分子机制。整体上,岩藻糖在细胞应激条件下,除了维持蛋白质折叠质量控制外,还把 UPR 从“损伤—促凋亡模式”拉回到“适应—恢复稳态模式”,进而稳住线粒体与关键糖蛋白的功能,为抗衰提供了蛋白稳态和细胞稳态。
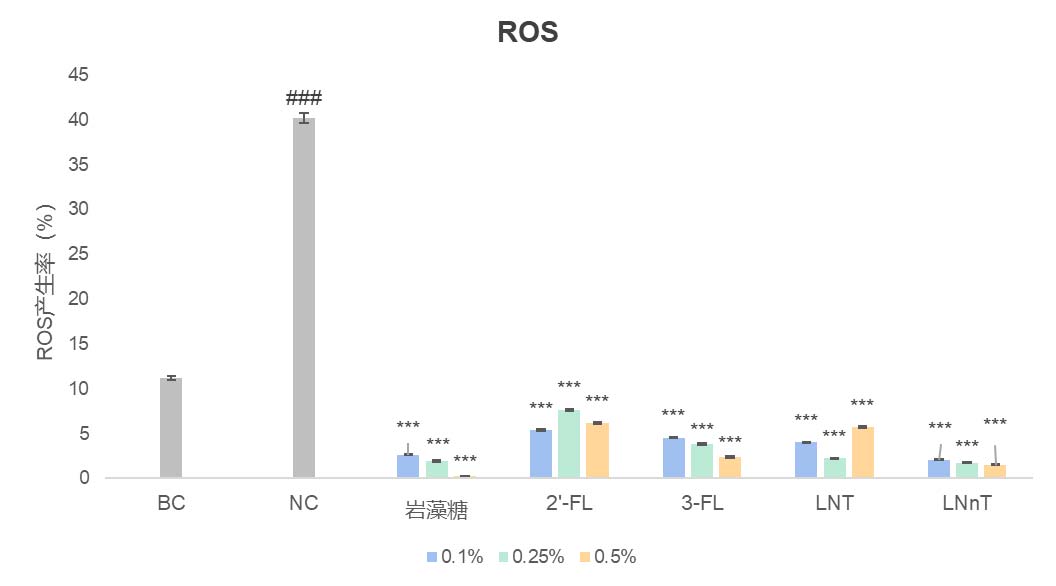
图5 岩藻糖显著降低LPS刺激的成纤维细胞的ROS
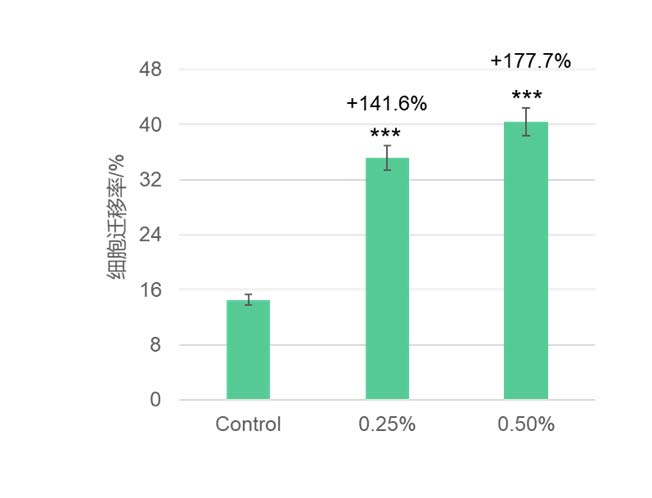
图6 岩藻糖显著提升角质形成细胞的迁移率
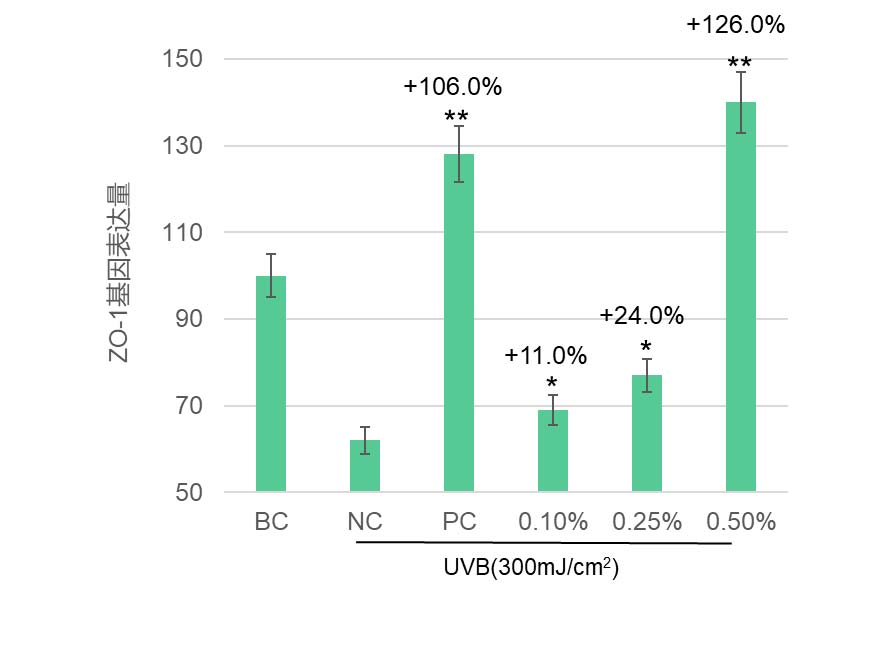
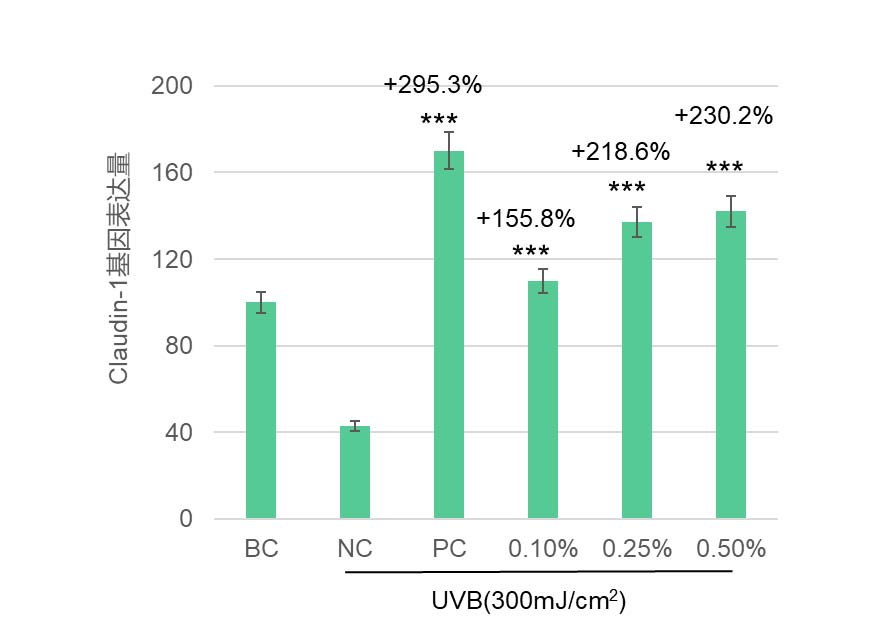
图7 岩藻糖显著提升角质形成细胞因UV受损的屏障蛋白表达
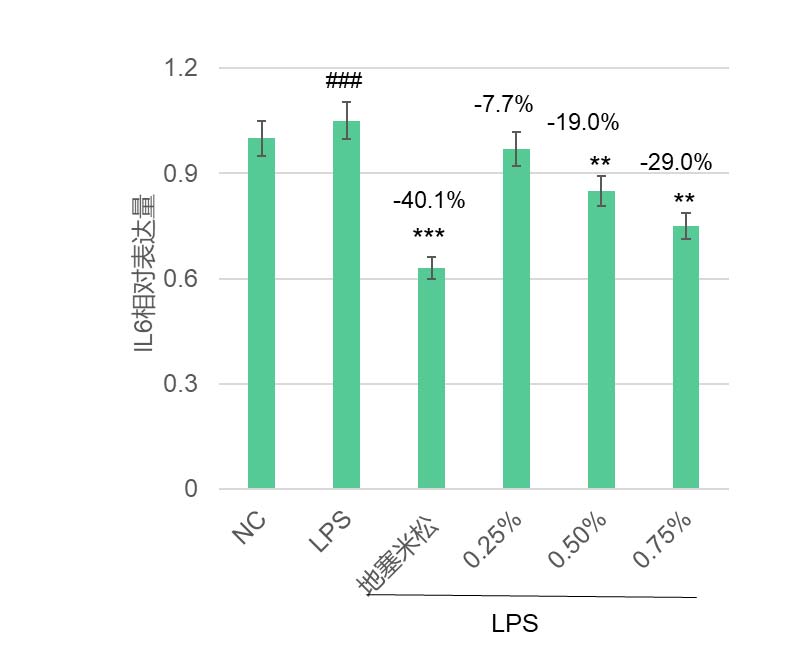
图8 岩藻糖抑制巨噬细胞促炎因子的表达

麦角硫因对于线粒体功能的保护和提升体现在三个方面:
• 靶向积累:麦角硫因通过OCTN1转运体主动摄取并富集于线粒体(Liu & Tang, 2023)。
• 抗氧化防御:麦角硫因调节ROS水平,维持氧化还原平衡,保护和维持线粒体功能(Liu & Tang, 2023)。
• 能量代谢保护和提升:麦角硫因增强线粒体膜电位;提高线粒体复合体I、II和IV的活性,从而提升呼吸链复合体功能,并促进能量代谢;增加线粒体与ATP生成相关的耗氧速率,促进了有氧呼吸的转化效率 (华熙内部数据)。
华熙生物通过D-gal 诱导的皮肤成纤维细胞衰老模型,发现衰老细胞经过麦角硫因的抗氧化防御和能量代谢提升作用,不仅增加了抗氧化酶和抗氧化因子的表达(图9和图10),恢复了衰老细胞的抗氧化能力,而且有效逆转了细胞的衰老表型,衰老相关基因 p21的高表达得到了相应的逆转和下降(图11)。在衰老细胞的胞内功能稳态得到恢复的同时,细胞外基质稳态也得到了重建,比如与基质降解相关的基质金属蛋白酶(MMPs)表达明显得到了抑制(图12), I 型胶原的合成也恢复了正常状态(图13)。
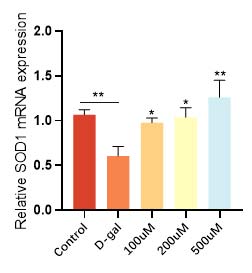
图9 麦角硫因升高衰老细胞抗氧化酶的表达水平
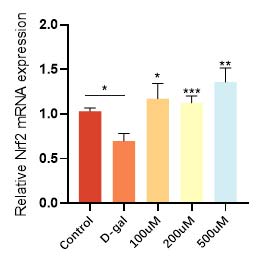
图10 麦角硫因升高衰老细胞抗氧化因子的表达水平
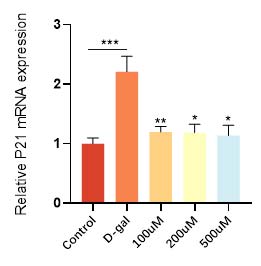
图11 麦角硫因降低衰老细胞p21的表达
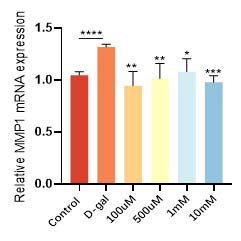
图12 麦角硫因降低衰老细胞基质金属蛋白酶的表达水平
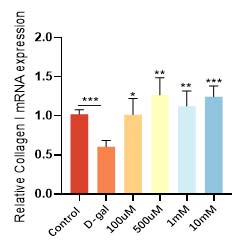
图13 麦角硫因升高I型胶原表达水平

岩藻糖和麦角硫因在皮肤细胞抗衰过程中形成了互补而协同的作用。
岩藻糖通过上调 FUT2 介导的岩藻糖基化,稳定内质网分子伴侣 HYOU1,从而维持蛋白质折叠质量控制与UPR平衡,避免促凋亡信号的激活,减轻氧化应激并保护线粒体功能。这使细胞在应激环境下能够将UPR从“损伤—凋亡模式”转向“适应—恢复稳态模式”,进而稳定线粒体膜电位、抑制促凋亡因子并提升抗凋亡能力。
与此互补,麦角硫因通过 OCTN1 特异性富集于线粒体,直接在能量代谢层面发挥保护作用:它清除过量ROS,提升复合体 I、II 和 IV 的活性,增强膜电位和ATP生成效率,从而全面恢复和提升线粒体的能量供给能力。实验显示,EGT 不仅增强了衰老细胞的抗氧化酶和抗氧化因子的表达,还逆转了 p21 等衰老标志基因的异常上调,抑制基质降解酶 MMPs 的过度表达,并恢复 I 型胶原合成。
由此可见,岩藻糖主要通过”内质网稳态—UPR—蛋白折叠“路径在内质网的层面提供蛋白质折叠和细胞稳态保护,而麦角硫因则通过”线粒体靶向—ROS调节—能量代谢提升“路径在线粒体层面提供能量代谢保护并维持氧化还原平衡支持。两者通过线粒体相关内质网膜(MAM) 紧密协作,协调能量代谢与应激应答,共同维护细胞稳态,提升对应激的应答能力,改善线粒体功能,抑制衰老表型,最终可在皮肤细胞层面实现了抗衰老的协同作用。

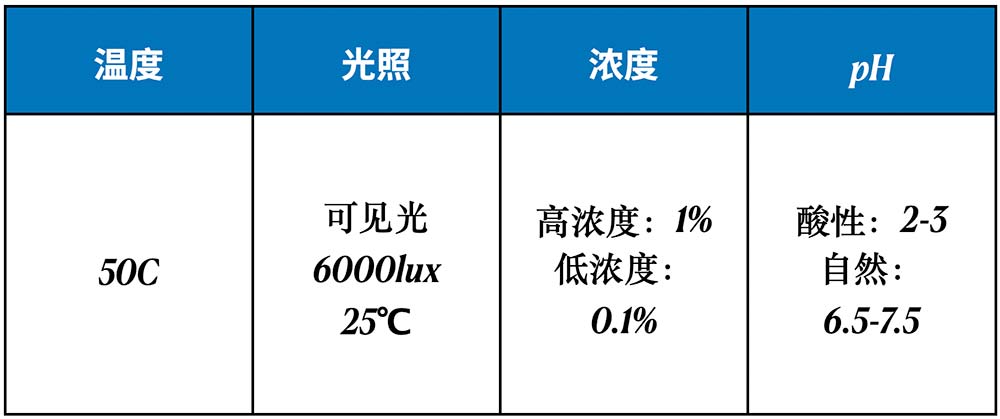
华熙生物比较了华熙麦角硫因和竞品公司麦角硫因之间,在稳定性上的差别。稳定性的条件选取了温度、光照、浓度和pH等四个与护肤品配方存储和使用相关的维度(表1),进行挑战实验。
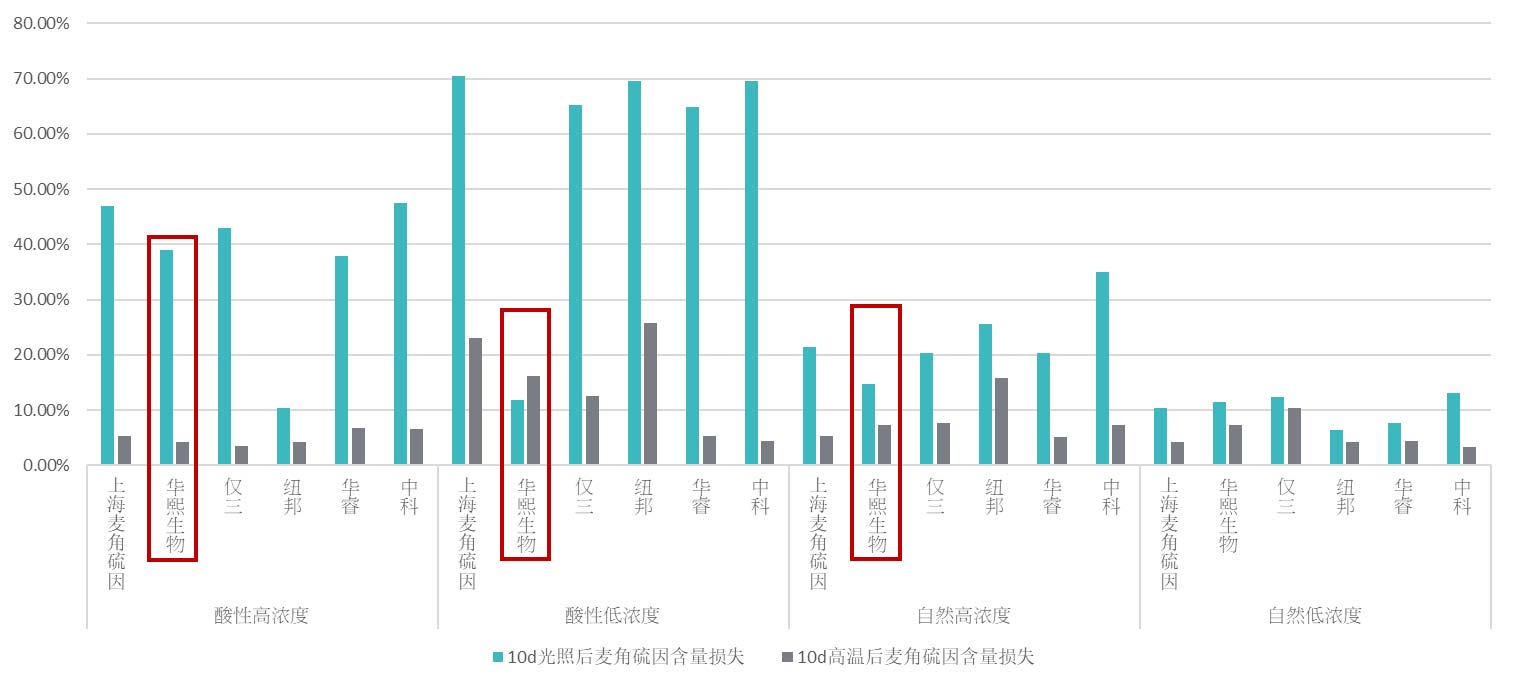
实验发现麦角硫因在0.1%浓度和酸性条件下,竞品麦角硫因极易受到光照的影响而降解(图14),比如上海麦角硫因、纽邦麦角硫因和中科麦角硫因的降解率达到70%,仅三和华睿降解率也在65%;而华熙麦角硫因的降解率仅为10%左右。由于0.1%正好是麦角硫因在护肤配方中的使用区间,且驻留型护肤配方的pH基本都在6以下,属于酸性区间,因此低浓度酸性条件下的光照稳定既可以保证麦角硫因在产品中的存储稳定性,又可以保证使用时麦角硫因免受日光的降解,极大提升产品的实际功效。
本文作者为孙欣博士。Bioπ中国美肤科学传播平台发布本文只是为了更多的信息参考,不代表任何有倾向性的投资意见或市场暗示。

THE END






指导单位:中国香料香精化妆品工业协会
受托运营:中国香妆融媒体
京ICP备2025131973号