
封面报道
Inflammaging(炎症性衰老)与成纤维细胞:
重新定义皮肤抗衰的医学核心
2026年05月25日 12:04
提起皮肤衰老,我们最先想到的总是细纹横生、面部松弛、肤色暗沉等表观的现象,并下意识把这些表现归咎于胶原蛋白的流失、保湿的不足或光损伤的累积这些偏微观的现象。然而,从皮肤生物学的角度来看,这些可见表现和微观现象都属于结果,而非真正的起点:几乎所有皮肤衰老的进程,都伴随着一场无声无息、持续不退的慢性低度炎症。这场炎症没有痘痘红肿刺痛的剧烈反应,没有晒伤后的灼热脱皮,它隐藏在真皮层深处,悄无声息却从未停歇,让皮肤看似稳定,却始终停留在“尚未完全结束的修复过程”中。于是在这一背景下,研究学者提出了 inflammaging ——一种由慢性低度炎症驱动的组织衰老模式[1-3]。
它改变了我们理解衰老与炎症的关系:皮肤并不是先老化,然后出现炎症;也不是某几次炎症导致了衰老,而是——皮肤在一生不断的微损伤与修复循环中,逐渐形成了一种无法被关闭的低度炎症背景,而衰老,是这种长期无休止炎症的组织结果[4]。
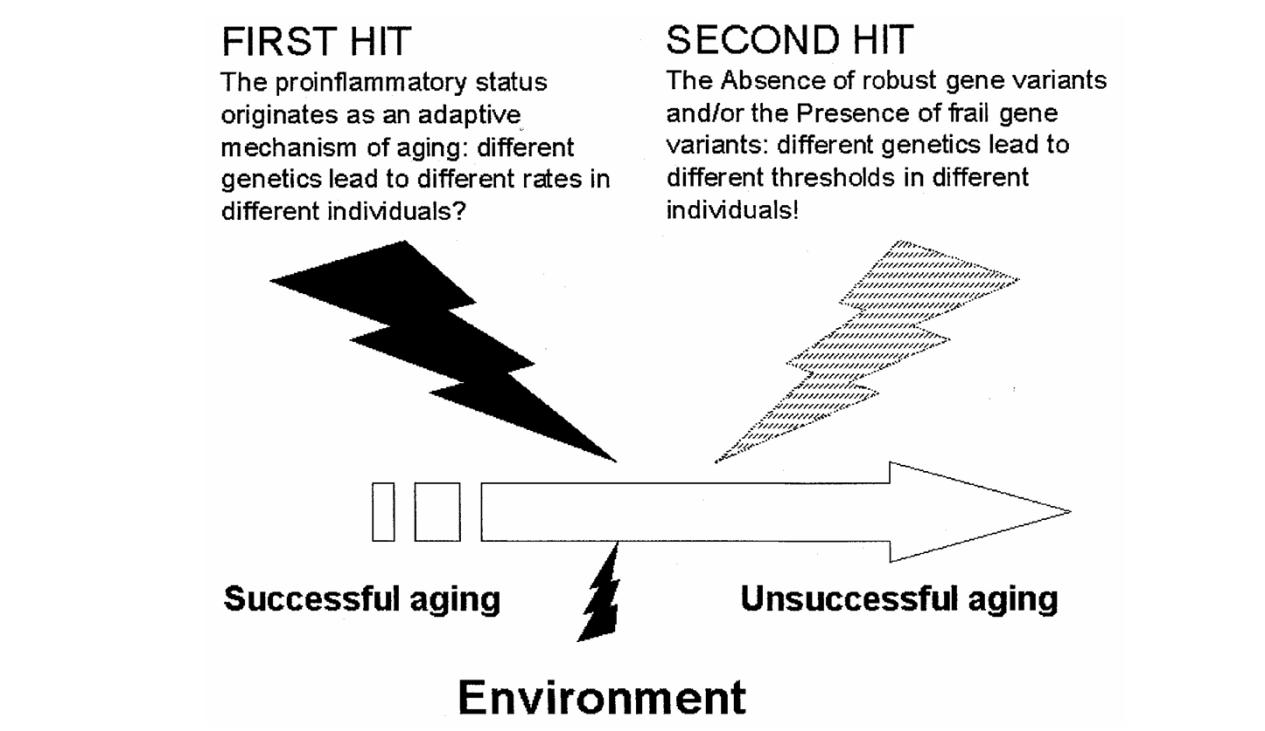
图1 炎症衰老的双重打击学说(PMID:10911963)
于是,一个更核心的问题出现了:如果炎症本应是短暂、可终止的生理过程,那么——为什么在衰老组织中,炎症停止的能力会逐渐丧失?
要回答这个问题,我们必须把视角从免疫细胞本身移开,重新审视长期被认为只是“结构支架”的组织细胞——
成纤维细胞,以及它所塑造的皮肤微环境。

皮肤作为人体最大的独立器官,其慢性炎症状态不同于内脏等组织的急性炎症反应,而是长期维持低度持续状态,这一特点与成纤维细胞功能改变及组织微环境重塑密切相关[5]。

成纤维细胞是真皮层的核心功能细胞,也是皮肤炎症的核心调控者,它的状态直接决定皮肤是否处于慢性炎症中:
年轻状态下,成纤维细胞活力旺盛,不仅负责合成胶原蛋白、弹性纤维、透明质酸,撑起皮肤饱满度,还能主动分泌抗炎因子,及时调整皮肤局部轻微炎症,维持微环境稳态,此时炎症只会短暂出现,快速消退。
而随着衰老启动,成纤维细胞会逐渐进入衰老僵尸状态,失去分裂和修复能力,并开启异常分泌模式,形成衰老相关分泌表型(SASP)——持续释放白细胞介素-6(IL-6)、肿瘤坏死因子-α(TNF-α)等促炎因子,同时大量分泌基质金属蛋白酶(MMPs),主动分解健康的胶原和弹性纤维[6–8]。
这种转变让成纤维细胞“倒戈”,从原本的抗炎修护者,兼而成为了慢性炎症的持续制造者,源源不断向皮肤组织释放炎症信号,让低度炎症再也无法停止。
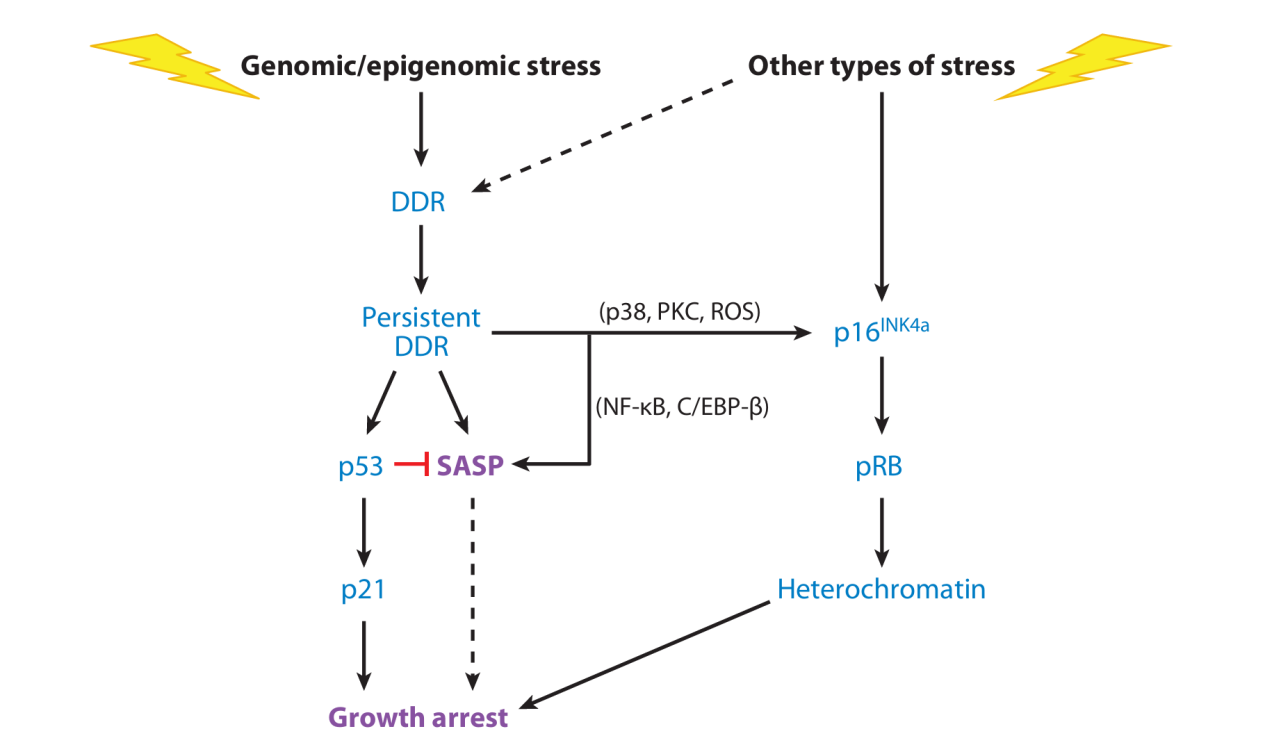
图2 衰老生长停滞和衰老相关分泌表型(SASP)的调控(PMID:23140366)

与此同时,皮肤的组织微环境,进一步放大了这种慢性炎症,也是它区别于其他组织的关键:
真皮层的细胞外基质(ECM),原本是传递修复信号、阻隔炎症扩散的屏障,但随着成纤维细胞衰老、基质被分解破损,这个网状结构变得松散塌陷,无法正常传递抗炎信号,反而会持续激活皮肤内的免疫细胞,让免疫细胞持续释放炎症因子[9,10]。
再加上皮肤表层屏障与微生态的联动失衡,破损的屏障无法阻挡外界微小刺激,失衡的微生态就会进一步诱发免疫反应[11],最终形成“衰老细胞→促炎因子释放→微环境恶化→更多细胞衰老”的死循环,让慢性炎症在皮肤组织内长期扎根,这就是Inflammaging的核心本质。
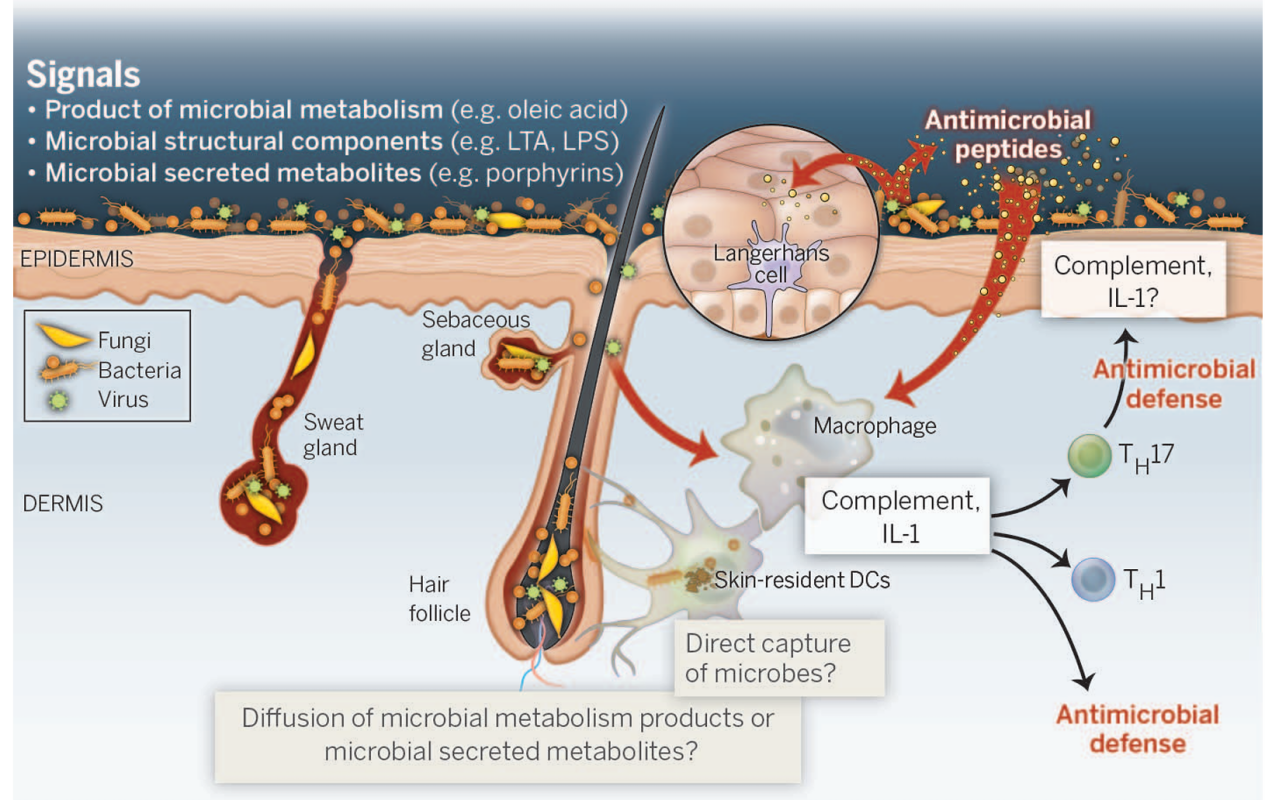
图3 微生物群与皮肤免疫系统之间的相互作用(PMID:25414304)

当炎症无法顺利终止的源头被发现后,一个现象变得格外清晰:慢性低度炎症几乎存在于所有衰老组织中,但皮肤是最容易观察到的衰老器官。
与肝脏、心脏或大脑不同,皮肤并不是一个被保护在体内环境中的器官。它是人体唯一长期直接暴露于外界的组织,也是机体最大的免疫界面。紫外线、空气污染、机械摩擦、微生物群变化、温度波动以及氧化压力,每时每刻都在不断触发皮肤的防御机制。
从免疫学角度看,皮肤始终处在一种“准备回应”的状态[12]。在年轻皮肤中,这种高警戒状态并不会带来问题。轻微损伤引发短暂炎症,随后迅速被关闭,组织完成修复并恢复稳态。然而随着年龄增长,皮肤逐渐失去这种精确的开关能力。
首先发生改变的是屏障系统。角质层更新速度下降,脂质结构紊乱,经皮水分流失增加,使外界刺激更容易进入组织内部。对机体而言,这意味着炎症触发阈值被持续降低——原本不需要反应的轻微信号,也可能被识别为损伤。
与此同时,紫外线长期累积造成的DNA损伤也在不断增加。研究表明[13],无法完全修复的DNA损伤会诱导细胞进入衰老状态,而衰老细胞并非简单“失去功能”,它们会持续分泌炎症因子、蛋白酶及趋化信号,这些分子进一步激活周围细胞,使局部组织长期处于低度炎症环境。于是,这些外界刺激日复一日的累积,成为皮肤慢性炎症的持续燃料,这是其他组织器官没有的致炎压力。
再加上上文中提到的,成纤维细胞决定炎症细胞在组织中的停留方式,随着年龄增长,它们持续表达趋化因子与存活信号,使免疫细胞更容易被保留在局部,而非完成任务后离开。
Buckley 等[14]研究提出,组织基质细胞具有“位置记忆(positional memory)”,不同部位的成纤维细胞会维持特定的炎症微环境。在衰老皮肤中,这种记忆可能被慢性刺激重新编程,使真皮层逐渐形成一种稳定的炎症生态位。
因此,皮肤之所以容易进入 inflammaging,并不仅因为外界刺激多,而是因为三个层面的因素同时发生改变:
• 持续暴露使炎症不断被启动;
• 细胞衰老使炎症信号持续被释放;
• 组织微环境改变使炎症难以终止。
炎症从一次事件,转变为一种组织状态。这也解释了临床中常见却难以归类的现象:许多老年皮肤并无明显炎症性疾病,却长期表现为干燥、敏感、修复缓慢与反复刺激反应。这些表现并非单一疾病,而是皮肤整体免疫—结构系统进入慢性炎症稳态后的结果。


当我们把皮肤衰老重新理解为一种长期慢性低度炎症状态时,抗衰老的临床目标也会随之发生改变。过去的治疗逻辑往往集中在“刺激以及再生”,通过能量设备、化学剥脱或高活性成分诱导组织修复反应,以获得胶原新生与结构重建,这种情况下刺激的“量”往往难以控制,刺激不足,则只是活性物的直接补充,皮肤并不能真正吸收利用;刺激过度,则会对皮肤造成伤害,得不偿失。然而在 inflammaging 背景策略下:衰老皮肤真正缺乏的,并不在于外在的刺激和补充,而是针对于皮肤内部的稳定。换句话说,许多熟龄肌、敏感肌或炎症后老化皮肤,并非修复能力不足,而是长期处于无法关闭的炎症状态中。
因此,现代皮肤科抗衰老策略开始从“以刺激手段促进皮肤自身修复”,逐渐转向更温和的:降低炎症负荷 → 重建微环境 → 恢复组织稳态。

遵循皮肤科温和修护原则,选用循证成分护肤品,长期阻断低度炎症、保护成纤维细胞,同时作为医美后基础修护方案:
• 靶向抗炎:选用皮肤科认可的温和抗炎成分,抑制促炎因子、阻断胶原分解,推荐高纯度油橄榄叶提取物、甘草酸二钾、泛醇、红没药醇,无激素无依赖,长期改善暗沉、敏感、隐性炎症。
• 激活成纤维细胞:用临床验证的修护抗衰成分,唤醒衰老细胞、促胶原再生,兼顾抗炎,核心选玻色因、医用重组类人胶原、温和胜肽,避开高浓度酸、视黄醇等刺激性成分,避免加重炎症应激。
• 屏障修复:选用含神经酰胺、角鲨烷、益生元的医用修护产品,加固皮肤屏障,切断外界致炎刺激入侵路径,改善炎症后色素沉着与反复敏感。
• 日常防晒:紫外线是光致炎症核心诱因,日常优先选氧化锌、二氧化钛为主的医用物理防晒,搭配硬防晒,从源头阻断光诱发慢性炎症,保护真皮层细胞。

针对中重度炎性衰老(松弛、顽固色斑、胶原大量流失),通过无创/微创医美项目,直达真皮层起效:
• 光电抗炎抗衰:强脉冲光(光子嫩肤)可封闭泛红、抑制浅层炎症、改善色沉,同时促胶原再生;黄金射频微针、热玛吉等射频项目,通过热能激活成纤维细胞,修复基质、调节免疫微环境,抗炎抗衰同步。
• 中胚层疗法:通过水光、微针导入医用抗炎修护精华(透明质酸、抗炎多肽、生长因子),直接作用于真皮层,快速平息慢性炎症,修复衰老成纤维细胞,改善干燥、细纹、松弛。
• 抗氧化抗炎导入:院线富勒烯、谷胱甘肽导入,清除自由基、减轻氧化炎症,减少衰老细胞SASP促炎因子分泌,辅助提升抗衰效果。

在 inflammaging 模型下,一个常被忽视的事实是:医美效果,很大程度取决于术后炎症是否被正确终止。过度刺激或错误护理,可能使治疗本身成为新的慢性炎症来源。
因此,术后管理应遵循三个原则:
• 减少二次刺激
避免频繁项目叠加、过度清洁或功效叠加。
• 优先抗炎修护
术后早期重点在抑制炎症,而非继续抗衰。
• 延长恢复窗口
给予组织足够时间完成真正修复,而非不断启动新一轮损伤。


现代抗衰的核心正在发生转移——从“刺激兼顾修护的手段换取再生”,转向更温和的“降低慢性炎症负荷,恢复组织稳态”。
无论是日常医用护肤、院线轻医美,还是术后维稳管理,其真正目标并非单一改善外观,而是在不同干预层级中持续调节皮肤微环境,使皮肤细胞,如成纤维细胞重新具备修复与响应能力。
当炎症被有效控制,皮肤所表现出的改变,往往不只是皱纹减少或紧致提升,而是一种更稳定的生理状态:耐受性增强、恢复周期缩短、衰老速度减缓。
从这一角度看,抗衰老不再是对抗时间的行为,而是一项关于炎症管理与组织功能维护的长期医学过程。

参考文献:
1. Franceschi C, Bonafè M, Valensin S, et al. Inflamm-aging: an evolutionary perspective on immunosenescence. Ann N Y Acad Sci. 2000;908:244–254.
2. Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014;69(Suppl 1):S4–S9.
3. Pilkington SM, Bulfone-Paus S, Griffiths CEM, et al. Inflammaging and the skin. J Invest Dermatol. 2021;141(4S):1087–1095.Fulop T. Gerontology, 2019.
4. Salminen A, Kaarniranta K, Kauppinen A. Inflammaging: disturbed interplay between autophagy and inflammasomes. Aging (Albany NY). 2012;4(3):166–175.
5. Quan T, Fisher GJ. Role of age-associated alterations of the dermal extracellular matrix microenvironment in human skin aging. Gerontology. 2015;61(5):427–434.
6. Coppé JP, Patil CK, Rodier F, et al. Senescence-associated secretory phenotype reveals cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6(12):2853–2868.
7. Campisi J. Aging, cellular senescence, and cancer. Annu Rev Physiol. 2013;75:685–705.
8. Quan T, Qin Z, Xia W, et al. Matrix-degrading metalloproteinases in photoaging. J Investig Dermatol Symp Proc. 2009;14(1):20–24.
9. Quan T, Fisher GJ. Role of age-associated alterations of the dermal extracellular matrix microenvironment in human skin aging. Gerontology. 2015;61(5):427–434.
10. Kaur A, Webster MR, Weeraratna AT. In the Wnt-er of life: Wnt signalling in melanoma and ageing. Br J Cancer. 2016;115(2):127–133.
11. Belkaid Y, Segre JA. Dialogue between skin microbiota and immunity. Science. 2014;346(6212):954–959.
12. Di Meglio P, Perera GK, Nestle FO. The multitasking organ: recent insights into skin immune function. Immunity. 2011;35(6):857–869.
13. Rodier F, Campisi J. Four faces of cellular senescence. J Cell Biol. 2011;192(4):547–556.
14. Buckley CD, Barone F, Nayar S, et al. Stromal cells in chronic inflammation and tertiary lymphoid organ formation. Annu Rev Immunol. 2015;33:715–745.
本文作者为医学博士、浙江大学医学院附属第二医院皮肤科副主任医师戴茹,戴茹长期从事皮肤科临床、教学与科研工作,主攻皮肤干细胞与毛囊再生、化妆品无创评价以及光电治疗与面部年轻化相关领域。Bioπ中国美肤科学传播平台发布本文只是为了更多的信息参考,不代表任何有倾向性的投资意见或市场暗示。

THE END
























京公网安备11010502058609号