
谈糖不色变
不只有胶原蛋白:
ECM的完整成分图谱
2026年06月09日 09:10
我们对ECM的认知,某种程度上像极了盲人摸象。胶原蛋白当然是真实的,而且重要。它是那条最显眼的“象腿”,支撑起了组织的基本力学框架。但问题在于,天然细胞外基质(ECM)从来不是“胶原蛋白的同义词”。
更准确地说,它是一个由胶原蛋白、弹性蛋白、糖蛋白(特指纤连蛋白、层粘连蛋白等非胶原糖蛋白)、蛋白聚糖等多种结构性分子共同组成的复杂体系[1]。胶原蛋白只是其中的骨架,骨架之外,还有让它真正“活起来”的其他部分。
这篇文章想做的,不是否定胶原,而是把那些长期被忽略、却同样决定ECM能否作为完整功能系统运转的部分介绍给你。

胶原蛋白不是一种蛋白,而是一个由28种成员组成的庞大家族,并且不同成员可以形成截然不同的超分子结构:有的形成纤维,有的形成网络,有的兼具蛋白聚糖特征[2],ECM的结构完整性正是建立在这个家族的协同分工之上。
其中,I型提供拉伸强度,III型赋予柔韧,这两位“明星成员”早已是行业常识。但胶原家族中,还有大量与皮肤功能直接相关、却很少被谈及的成员。
比如IV型胶原不形成典型的纤维,而是自组装成网状结构,构筑基底膜的核心骨架[2]。在皮肤中,基底膜是上皮与真皮之间的关键界面,营养滤过、细胞迁移引导、表皮干细胞微环境维持,都依赖这张网络。研究显示,IV型胶原在真皮表皮交界处的表达随年龄增长而逐步减弱,伴随表皮嵴(rete ridges)结构的进行性消失[3]。这是皮肤老化过程中基底膜变薄、真皮表皮交界趋于平坦的结构基础之一。
此外,VI型胶原形成串珠状微纤维,在弹性纤维与细胞基底膜之间搭建物理连接[2];VII型胶原组装成锚定纤维,将表皮固定在真皮上,缺失即可导致表皮与真皮分离[2];XV型和XVIII型胶原的C端非胶原结构域在被蛋白酶切割后释放具有抗血管生成活性的片段,内皮抑素(endostatin)即来自XVIII型胶原[2]。


胶原家族负责让组织“撑得住”,弹性蛋白负责让组织“弹得回”。
弹性蛋白是脊椎动物相对晚近的进化产物,它的出现与闭合循环系统及更高水平的弹性回缩需求密切相关[4]。皮肤中每一次表情、触碰、牵拉之后恢复原状,依赖的正是弹性蛋白赋予的可逆形变能力。
弹性蛋白最特殊的生物学特征,在于它的极低更新速率和成年后几乎不可再生的特性。成熟弹性纤维并不是单一弹性蛋白,而是弹性蛋白与微纤维共同构成的复合结构。更关键的是,原弹性蛋白(tropoelastin) 在细胞外会发生高度交联,超过 80% 的赖氨酸残基参与形成共价交联;正是这种高密度交联,赋予了它极强的回弹能力,也带来了极长寿命。成体后弹性蛋白的新合成几乎停止,每年降解更新不到全身总量的1%[4]。
这意味着弹性纤维一旦损伤,成年机体很难真正重建。这正是瘢痕组织僵硬无弹性的结构基础之一,也是皮肤老化过程中弹性回缩能力持续下降的根本原因。因此,对 ECM 类材料而言,是否保留了弹性纤维相关线索,至少应成为评价“是否更接近天然 ECM”的一个维度。


胶原和弹性蛋白解决的是“撑得住”和“弹得回”。但仅有结构骨架还不够,细胞需要知道自己身处何处、该做什么。ECM之所以能回答这些问题,是因为它内部存在一套组织逻辑,而这套逻辑,很大一部分由ECM糖蛋白承担。
而在这些分子中,纤连蛋白(fibronectin)和层粘连蛋白(laminin)尤其关键。它们的共同特征是,都携带可被细胞表面受体识别的结合位点。RGD序列是纤连蛋白等ECM配体中最经典的例子之一,但并非所有ECM糖蛋白都依赖RGD完成识别[5]。不同糖蛋白通过不同结构域与不同整合素亚型结合,触发不同的细胞内信号通路。换句话说,ECM糖蛋白不是在笼统地告诉细胞“你可以粘上来了”,而是在更精确地告诉细胞“你是谁、你在哪、你该往哪走”。
但糖蛋白的功能远不止信号识别。纤连蛋白是ECM组装的核心组织者,多种基质蛋白,包括胶原、fibrillin、LTBP等的沉积,都依赖于纤连蛋白纤维网络的预先建立[6]。层粘连蛋白则是基底膜自组装的重要起始组织者,层粘连蛋白网络通常先行建立,并为IV型胶原网络在正确位置形成提供框架[6]。此外,纤连蛋白上的FN3结构域可以同时结合整合素和VEGF等生长因子,将黏附信号和生长因子信号在同一个分子上编排到一起,让细胞同时接收到多重指令[6]。
ECM糖蛋白既是细胞与基质对话的信号接口,也是基质自身组装的骨干。
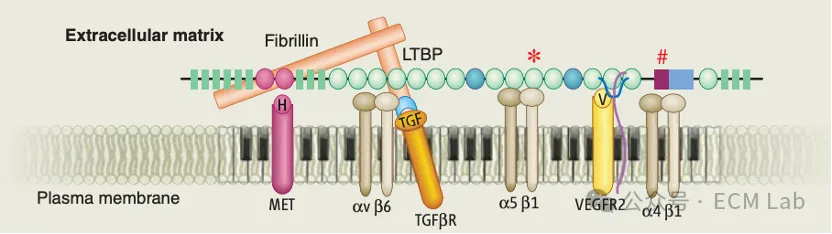
图1|ECM糖蛋白既是信号接口,也是基质组装的组织者:
来源:引自 Hynes RO. The extracellular matrix: not just pretty fibrils. Science, 2009, 326(5957): 1216–1219, Fig. 3.

如果糖蛋白决定“细胞怎么接触 ECM”,那么糖胺聚糖(GAG)和蛋白聚糖决定的,是细胞在什么时候、什么位置,收到什么信号。
GAG是ECM中一大类长链多糖分子,其中透明质酸(HA)最广为人知。它不经硫酸化修饰,主要依靠极强的吸水能力为组织提供水合、黏弹性和体积支撑。相比之下,真正让ECM具备精细信号调控能力的,往往是另一组成员,即硫酸化GAG及其所在的蛋白聚糖体系。
其中最具代表性的是硫酸乙酰肝素(HS)。HS并不是一条结构完全固定的糖链,而是一种高度异质性的分子,不同修饰模式会改变它对不同生长因子的结合偏好[7]。这意味着它并不是一视同仁地把所有因子都“粘”在表面,而更像是在筛选、分配和呈递不同的信号分子。虽然ECM成分并不包括生长因子,但它可以通过蛋白聚糖网络结合多种生长因子,在局部完成富集、保护和条件触发释放——从而精细调控这些信号的时空分布[7]。
这种调控逻辑在TGF-β身上体现得尤其充分。TGF-β通常并不是以自由活性形式存在,而是以潜伏复合物形式被整合进ECM中,锚定于纤连蛋白和微纤维等结构[6][7]。只有在蛋白酶切割、构象变化或细胞牵拉等特定条件下,它才会被局部激活释放。正是这套系统的存在,让ECM具备了对生长因子进行时空编辑的能力。它不只是储存信号,而是决定信号何时、何地、以何种浓度被细胞接收。
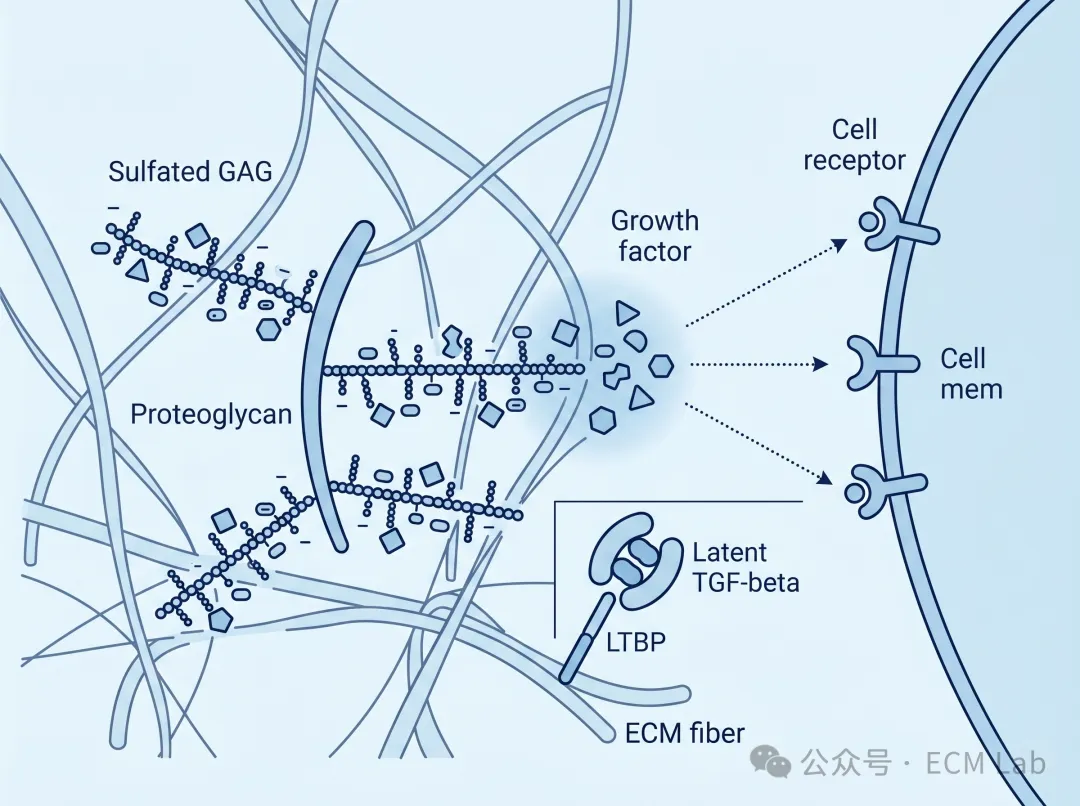
图2|硫酸化GAG与蛋白聚糖通过锚定、富集与局部呈递组织信号:在 ECM 中,硫酸化 GAG 链并不是被动支撑成分,而是通过与生长因子结合,形成局部信号储库与呈递界面;邻近细胞接收到的,不再是自由扩散的背景信号,而是被 ECM 组织过、具有空间指向性的微环境信息。

天然ECM的价值
不在“某个成分”,而在“整套组织逻辑”
回到开头那个问题:当我们说 ECM 时,我们到底在说什么?
答案显然不只是胶原。
我们说的是一个由胶原家族提供结构框架、由弹性系统赋予回弹能力、由糖蛋白组织装配并建立识别界面、由 GAG/蛋白聚糖管理生长因子时空分布的复杂系统。它不是一堆成分的相加,而是一套被组织起来的“环境逻辑”。
所以,真正值得追问的,从来不只是“ECM里有没有胶原”,而是: ECM 赖以发挥作用的那套结构、弹性、界面、信号与组织逻辑,是否被完整地认知。
而这,也正是下一篇最值得继续讲下去的部分: 决定 ECM 最终“说什么”的,不只是它含有什么,更是这些成分在三维空间里如何被组织到一起。
参考文献
[1] Naba A, Clauser KR, Hoersch S, Liu H, Carr SA, Hynes RO. The matrisome: in silico definition and in vivo characterization by proteomics of normal and tumor extracellular matrices. Molecular & Cellular Proteomics, 2012, 11(4): M111.014647.
[2] Ricard-Blum S. The collagen family. Cold Spring Harbor Perspectives in Biology, 2011, 3(1): a004978.
[3] Roig-Rosello E, Rousselle P. The human epidermal basement membrane: a shaped and cell instructive platform that aging slowly alters. Biomolecules, 2020, 10(12): 1607.
[4] Wagenseil JE, Mecham RP. Vascular extracellular matrix and arterial mechanics. Physiological Reviews, 2009, 89(3): 957–989.
[5] Humphries JD, Byron A, Humphries MJ. Integrin ligands at a glance. Journal of Cell Science, 2006, 119(19): 3901–3903.
[6] Hynes RO. The extracellular matrix: not just pretty fibrils. Science, 2009, 326(5957): 1216–1219.
[7] Taipale J, Keski-Oja J. Growth factors in the extracellular matrix. FASEB Journal, 1997, 11(1): 51–59.
来源:ECM Lab微信公众号 Bioπ中国美肤科学传播平台发布本文只是为了更多的信息参考,不代表任何有倾向性的投资意见或市场暗示。

THE END
























京公网安备11010502058609号